- функція [ правити | правити код ]
- Мова і характеристики вокалізації [ правити | правити код ]
- діалекти [ правити | правити код ]
- Загальні характеристики [ правити | правити код ]
- Присвоєння і імітація [ правити | правити код ]
- нейрофізіологія [ правити | правити код ]
Вокалізація птахів - голосові звуків , що видаються птахами . В орнітології розділяється на довге спів і короткі голосові сигнали, різні за функціями - спів використовується переважно в статевій поведінці і для захисту території, голосові сигнали призначені переважно для комунікації, наприклад призову або повідомлення про небезпеку.
Більшість звуків, що видаються птахами, є мелодійним для людського вуха і вплинуло на розвиток музики.
У вокалізації виділяють спів і голосові сигнали, відмінність між якими ґрунтується на модуляції , Довжині і контексті звуків. Спів або пісня довга і більш складна і асоціюється з шлюбним і територіальним поведінкою, тоді як голосові сигнали або заклики виконують функції попередження або утримання зграї разом [1] [2] .
Спів найбільш розвинене у птахів загону воробьінообразних , Особливо підгрупи співочі птахи . Здебільшого спів властиво самцям , а не самкам , Хоча трапляються винятки. Спів найчастіше видається, коли птах сидить на якомусь субстраті, хоча деякі види здатні видавати його під час польоту . Деякі групи птахів практично мовчазні, видають лише механічні звуки, наприклад лелека , Лише клацають дзьобами . У деяких Манакін (Pіprіdae) самці розвинули кілька механізмів утворення таких звуків, включаючи стрекотіння, характерне для комах [3] .
Освіта звуків механічними засобами, на відміну від Сірінкс , Носить назву інструментальної музики (за визначенням Чарльза Дарвіна ) Або механічних звуків [4] і, в роботах сучасних авторів, Сонаце [5] . Термін Сонаце означає як акт освіти невокальних звуків, які утворюються з певним призначенням, і є комунікаційними сигналами, які утворюються неголосовими структурами, такими як дзьоб , Крила, хвіст і пір'я [5] .
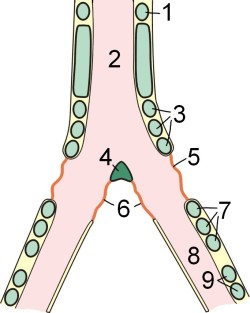
Схема будови Сірінкс
1 - остання вільна хрящове кільце трахеї; 2 - тимпанальной орган; 3 - перша група кілець Сірінкс; 4 - козелок; 5 - бічна тімпанальная мембрана; 6 - середня тімпанальная мембрана; 7 - друга група кілець Сірінкс; 8 - бронх; 9 - хрящові кільця бронха
Голосовим органом птахів є сірінкс . Це кісткова структура в місці біфуркації трахеї . На відміну від ссавців, у птахів відсутні голосові складки. Звук видається внаслідок вібрацій тимпанальной мембран (стінок Сірінкс) і козелка, що викликаються продуванням повітря через сірінкс [6] . Спеціальні м'язи здатні змінювати натяг мембран і діаметр просвіту бронхів, що призводить до зміни видаваного звуку.
Сиринкс і іноді повітряні мішки , Які оточують його, резонують у відповідь на вібрації, які створюються мембранами, повз яких проходить повітря при диханні. Птах контролює частоту звуку зміною натягу мембран. Так птиці контролює як частоту, так і гучність, змінюючи швидкість видиху. Птахи здатні незалежно контролювати обидві сторони трахеї, тому деякі види утворюють дві головні частоти одночасно.
функція [ правити | правити код ]
Прийнято вважати, що спів птахів розвинулося насамперед в результаті статевого відбору як елемент статевої поведінки , Зокрема залицяння і залучення самок самцями. Крім того, іншою важливою функцією співу є позначення території . За даними експериментів, якість вокалізації є індикатором пристосованості до умов навколишнього середовища [7] . Також, за даними експериментів, на характеристики і частоту співу можуть впливати паразити і захворювання, таким чином, вокализация є прямим індикатором здоров'я [8] [9] . Репертуар співу також є важливим індикатором пристосованості [10] [11] , Здатності самців привертати самок і позначати територію. Часто різні за функцією типи співу виробляються лише протягом певного сезону або в різні пори року, коли необхідно виконання певної функції, і лише протягом цього часу вони сприймаються іншими птахами. Наприклад, самець західного солов'я (Luscіnіa megarhynchos) видає спів, призначене для залучення самок, лише вночі (коли співають лише неспарені самці), а спів, призначене для позначення території, - майже виключно на протязі ранкового хору (коли співають всі самці) [12] .
Голосові сигнали використовуються насамперед для комунікації. Таке спілкування здійснюється як в межах одного виду, так і між видами. Загальні сигнали часто використовуються для залучення індивідуальних птахів до зграї. Ці голосові сигнали характеризуються широким спектром і різким початком і закінченням, а їх повторення, звичайне серед багатьох видів, як вважається, корисно для визначення розташування зграї. Сигнали попередження про небезпеку, на відміну від них, в більшості видів характеризуються високою частотою звуку, який ускладнює визначення положення птиці, яка видає такий сигнал [13] .
Часто птахи можуть дуже добре розрізняти голосові сигнали, що дозволяє їм розпізнавати один одного по голосу. Зокрема, багато птахів, що гніздяться в колоніях, таким чином розпізнають своїх пташенят [14] .
Багато птахів здатні подавати сигнали дуетом. Часом такі дуети настільки синхронізовані, що звучать як один голосовий сигнал. Такі сигнали називають антифону [15] . Дуетні сигнали відзначені в багатьох родинах птахів, включаючи Фазанові [16] , Саванних сорокопудів ( Malaconotidae ) [17] , тимелієві і деяких совоподібних [18] і попугаеобразних [19] . Сухопутні співочі птахи найчастіше роблять подібні сигнали в разі вторгнення чужинців на їх територію [20] , Пропонуючи роль таких сигналів в міжвидової конкуренції.
Деякі птахи здатні дуже добре імітувати голосові сигнали. У деяких птахів, таких як дронгові , Імітування сигналів може служити для формування многовидових зграй [21] .
Деякі печерні види, такі як гуахаро і салангани (роду Collocalia і Aerodramus ), Використовують звуки в діапазоні переважно від 2 до 5 кГц для ехолокації в темряві печер. [22] [23] [24] .
Мова і характеристики вокалізації [ правити | правити код ]
Мова птахів протягом тривалого часу був темою міфів і легенд. Дуже давно було відомо, що голосові сигнали мають певне значення, яке відповідним чином інтерпретується слухачами. Домашні кури , Наприклад, мають різні сигнали у відповідь на наближення повітряних і наземних хижаків, і відповідним чином відповідають на них [25] [26] . Проте, мова , На додаток до окремих словами , Повинен мати і певні граматичні структури і правила. Дослідження подібних структур у птахів є досить важкими через великої кількості можливих інтерпретацій. В одному дослідженні, проте, дослідникам вдалося продемонструвати здатність папуг до утворення граматичних структур, включаючи наявність таких понять, як іменник , дієслово і прикметник [27] . Дослідження голосових сигналів шпаків також показали наявність рекурсивних структур [28] .
Зазвичай при описі мови птахів мисливцями і натуралістами виділяють 5 основних типів звуків: заклик, пісня, територіальний сигнал, залицяння і тривога. Перші чотири представляють «базове» поведінку і подаються при відносній безпеці і спокої, тоді як останній означає присутність хижака або іншої загрози. В межах кожної категорії значення звуків залежать від модуляції голосу, рухи тіла і контексту [29] .
слух птахів може виходити за межі слуху людини, опускаючись у деяких видів як нижче 50 гц , Так і вище 20 кГц, з максимальною чутливістю між 1 і 5 кГц [30] [31] .
Діапазон частот голосових сигналів залежить від умов навколишнього середовища, перш за все наявних шумів. Зазвичай вузькі частотні діапазони, низькі частоти, низькочастотна модуляція і довга тривалість звуків і інтервалів між ними характерні для просторів з щільною рослинністю (де відбувається поглинання та відбиття звуків), тоді як високі частоти, широкі діапазони, високочастотна модуляція і короткі елементи сигналів характерні для відкритих просторів [32] [33] . Була також запропонована теорія, згідно з якою наявний частотний і часовий діапазон ділиться між різними птахами і їх видами, в результаті чого, при його обмеженні довжина і частотна ширина звукових сигналів зменшуються, цей ефект відомий під назвою «акустичної ніші» [34] . Птахи співають голосніше і на більш високих частотах в міських районах, де є значний низькочастотний шум [35] [36] .
діалекти [ правити | правити код ]
Вокалізація птахів навіть одного виду часто досить сильно відрізняється, утворюючи «діалекти». Ці діалекти можуть виникати як через різноманітність навколишнього середовища, так і внаслідок генетичного дрейфу , Хоча явище мало досліджено, вплив окремих факторів залишається невідомим навіть для добре досліджених видів [37] . Найкраще ці відмінності досліджені для співу під час шлюбного періоду. Проте, наслідки цього явища не є однаковими і істотно відрізняються в залежності від виду птахів.
Самки, які виросли під впливом одного діалекту, не відповідають або відповідають гірше на спів самця того ж виду, який володіє іншим діалектом, що було продемонстровано, наприклад, для білоголової зонотріхіі ( Zonotrichia leucophrys ) [38] [39] . З іншого боку, самки, які відбуваються з районів, де поширені кілька діалектів або діалекти різних підвидів, не виявляють такої переваги до одного діалекту [40] [41] .
Також був досліджений і відповідь територіальних самців на спів чужинців. Так, звичайно самці найсильніше відповідають на спів представників власного діалекту, слабкіше на представників свого виду з інших районів, і ще слабше на спів споріднених видів [42] , А самці, які поділяють більше пісень з сусідніми, краще охороняють свою територію [43] .
У зв'язку з виникненням діалектів часто розглядається питання їх впливу на видоутворення . Наприклад, це явище було продемонстровано в роботах по дослідженню в'юрків Дарвіна [44] . Інші роботи, проте, вказують на суперечливість даних з цього питання [45] [46] [37] .
Загальні характеристики [ правити | правити код ]
Спів птахів різних видів досить сильно відрізняється одне від іншого і часто є характерною ознакою виду. Саме спів часто є особливістю, яка перешкоджає змішання споріднених видів, генетично близьких для створення життєздатного потомства [47] . У сучасних дослідженнях спів характеризується за допомогою акустичної спектроскопії . Види сильно відрізняються за складністю співу і по числу типів пісень, яке може досягати 3000 у бурого пересмішника; у деяких видів навіть окремі особини відрізняються цією характеристикою. У декількох видів, таких як шпаки і пересмішники , Спів включає випадкові елементи, після успішної реєстрації протягом життя птиці в формі мімікрії або «привласнення» (через те, що птах використовує звуки, характерні для інших видів). Ще в 1773 році було встановлено, що в експериментах з вирощування пташенят птахами інших видів, коноплянка (Acanthіs cannabіna) була здатна вивчити спів жайворонка ( Alauda arvensis ) [48] . У багатьох видів здається, що хоча основна пісня і однакова для всіх представників виду, молоді птахи вивчають деякі деталі співу від своїх батьків, при цьому варіації накопичуються, формуючи «діалекти» [49] .
Зазвичай птиці вивчають пісні протягом початку життя, хоча певні особливості продовжують накопичуватися і пізніше, формуючи спів дорослої птиці. Зебровая амадина , популярний модельний організм для дослідження співу птахів, формує спів, яке нагадує доросле, приблизно після 20 діб після вилуплення. До віку 35 днів пташеня вже повністю вивчає доросле спів. Найбільш ранні пісні є досить «пластичними» або схильними до змін, і птиці треба приблизно 2-3 місяці для доведення співу до остаточної незмінною форми у статевозрілих птахів [50] .
Дослідження також вказують на те, що навчання співу є формою навчання, в якій беруть участь ділянки базальних гангліїв . Часто моделі навчання птахів використовуються в якості моделей вивчення мови людиною [51] . У деяких видів (наприклад, зеброва амадина ) Навчання обмежена першим роком життя, ці види називають «обмеженими за віком» або «закритими». Інші види, такі як канарейка , Здатні вивчати нові пісні навіть в статевозрілому віці, такі види називаються «відкритими» або «необмеженими за віком» [52] [53] .
Дослідники висловлювали припущення, що навчання пісням через широке культурне спілкування дозволяє освіту міжвидових діалектів, які допомагають птахам адаптуватися до різноманітного акустичному оточенню [54] .
Навчання птахів батьками було вперше продемонстровано в експериментах Вільяма Торпі 1954 року . Вирощені в ізоляції від самців свого виду птахи здатні співати, а їх спів за загальними рисами зазвичай нагадує спів дорослих птахів, проте, не має складних елементів і часто істотно відрізняється [55] . Таке спів часто не здатна привертати самок. Крім співу батьків, пташенятам також важливо чути своє власне спів протягом сенсоромоторной періоду. Птахи, які втратили слух до кристалізації співу, виробляють спів, яке істотно відрізняється від характерного для даного виду [56] .
Присвоєння і імітація [ правити | правити код ]
Багато птахи здатні переймати спів не тільки свого виду, але і інших, більш-менш споріднених видів. Так, пташенята багатьох видів, вирощені батьками споріднених видів, часто здатні розвивати спів, яке нагадує спів прийомних батьків, і в деяких випадках навіть привертати самок цього виду [44] . Інші птахи здатні переймати спів птахів інших видів, навіть коли вирощуються власними батьками. Кілька сотень видів по всьому світу здатні до такої імітації. Наприклад, назва пересмішник (Mіmus) було дано цього птаха саме за здатність копіювати звуки інших птахів і відтворювати їх. Іншим відомим видом, здатним до копіювання, є звичайного шпака (Sturnus vulgarіs), зокрема в Північній Америці , Куди ця птиця була завезена з Європи, він «передражнює» навіть пересмішника. У Європі та Британії звичайного шпака є найвідомішим імітатором співу інших птахів, який часто відтворює звуки таких птахів, як канюк звичайний (Buteo buteo), oriolus oriolus , Numenius arquata , сіра сова (Strіx aluco), качині і гуси . У деяких випадках ці птахи здатні імітувати голос немовляти або навіть звуки падаючих бомб під час Другої світової війни. За деякими даними, один шпак зімітував свисток футбольного судді, чим викликав непорозуміння під час матчу [57] .
Найбільш вражаючим і популярним серед людей прикладом імітації звуків птахами є імітація людської мови. відомо кілька хвилястих папужок , Які виросли в неволі, чий репертуар сягав 550 слів. Також хорошими здібностями для імітації мови людини володіють папуга жако (Psіttacus erіthacus), австралійські папуги, такі як какаду ( Cacatua galerita ) І південноамериканські амазони (Amazona). Олександр фон Гумбольдт під час дослідження Південної Америки описав випадок, коли йому вдалося почути від папуги «мертву мову» зниклого племені АТУР. В Європі випадки здатності імітувати голос людини були відомі серед деяких представників сімейства воронових , такі як галка (Corvus monedula), сорока (Pіca pіca) і ворон (Corvus corax) [58] .
Проте, точні причини такої імітації невідомі. Ймовірно, вони є ускладненнями власного співу, однак користь такого явища для птиці все ще залишається предметом досліджень.
Також існують випадки привласнення голосових сигналів, а не співу птахів. наприклад, толстоклювая еуфонія ( Euphonia laniirostris ) Часто видає сигнал загрози інших видів під час наближення потенційного хижака до її гнізда, залишаючись у безпеці. Подібна поведінка характерно і для сойки (Garrulus glandarіus) і червоноголової горихвістки ( Cossypha natalensis ). В інших випадках імітація використовується для заманювання жертви в пастку, так, димчастий лісової сокіл ( Micrastur mirandollei ) Здатний імітувати заклики про допомогу своїх жертв, а потім ловить птахів, які прилетіли у відповідь на заклик [59] .
нейрофізіологія [ правити | правити код ]
В контролі голосових сигналів беруть участь такі відділи головного мозку:
- Шлях до списку: складається з верхнього вокального центру (Hіgh vocal center or hyperstrіatum ventralіs pars caudalіs, HVC), ядра аркопілліума ( розривне ядро arcopіllіum, RA) і підпадає під язик, якраз до трахеї і сірінкс ( трахеосирінгеальний нерв) [60] [61] ;
- Передня частина переднього мозку, яка є для навчання: зробила з латеральної частини магноцеллюлярної ядра переднього нового смугастого тіла (бічна частина магноцелюлярного ядра переднього неостриатуму, LMAN, гомолога базових гангліїв ссавців ), Область X (користується базовим ганглієм) і дорсально-латерального відділення середнього таламуса (DLM).
- ↑ Ерліх, Пол Р., Девід С. Добкін і Дарріл Віє. "Пташині голоси" та "Вокальний розвиток" з нарисів Птахів Стенфорда (неопр.). Дата центрагу 9 вересня 2008 року. Читальний зал 7 квітня 2012 року.
- ↑ І. Р. Беме. Формування вокалізуючих горобіних птахів (Passeriformes) в онтогенезі. Сучасний стан проблем (Рос.) // Журнал загальної біології: журнал. - 2006. - Т. 67, № 4. - С. 268-279. - PMID 17022487 . синопсис
- ↑ Боствік, Кімберлі С. та Річард О. Пром. Охоплення птахів співає стригучим пір’ям крила (англ.) // Наука: журнал. - 2005. - Вип. 309, ні. 5735. - С. 736. - DOI : 10.1126 / наука.1111701 . - PMID 16051789 .
- ↑ Manson-Barr, P. and Pye, JD (1985). Механічні звуки. «Словник птахів» (ред. Б. Кемпбелл та Е. Лакк), стор 342-344. Стаффордшир: Пойсер.
- ↑ 1 2 Боствік, Кімберлі С. та Річард О. Пром. Високошвидкісний відео-аналіз розкриття крил у двох манакінових кладах (Pipridae: Aves) (Англ.) // Журнал експериментальної біології : Журнал. - Компанія біологів (Англ.), 2003. - Вип. 206. - С. 3693-3706. - DOI : 10.1242 / jeb.00598 . - PMID 12966061 .
- ↑ Безпосереднє спостереження за функцією м’язів шприца у птахів-пісень та папуг Оле Незбі Ларсен та Франц Голлер, Журнал експериментальної біології 205, 25-35 (2002)
- ↑ Прочитайте, AW та DM Weary. Сексуальний відбір та еволюція пташиної пісні: Тест гіпотези Гамільтона-Зука (Англ.) // Екологія поведінки та соціобіологія : журнал. - 1990. - Вип. 26, ні. 1. - С. 47—56. - DOI : 10.1007 / BF00174024 .
- ↑ Гарамшегі, LZ, AP Møller, János Török, Gábor Michl, Péter Peczely та Murielle Richard. Імунний виклик опосередковує голосовий зв’язок у перехожих птахів: експеримент (неопр.) // Поведінкова екологія. - 2004. - Т. 15, № 1. - С. 148—157. - DOI : 10.1093 / beheco / arg108 .
- ↑ Redpath, SM, Bridget M Appleby, Steve J Petty. Чи зраджують чоловічі хути навантаження на паразитів у Таун-Сови? (Англ.) // Журнал біології птахів : журнал. - 2000. - Вип. 31, вип. 4. - С. 457—462. - DOI : 10.1034 / j.1600-048X.2000.310404.x .
- ↑ Рейд, Дж. М., Пітер Арсез, Еліс ЛЕВ Кассіді, Сара М. Гіберт, Джеймс Н. М. Сміт, Філіп К. Стоддард, Емі Б. Марр та Лукас Ф. Келлер. Фітнес-корелянти розміру пісенного репертуару у вільноживих пісенних горобців ( Melospiza melodia ) (англ.) // Американський натураліст (англ.): журнал. - Університет Чикаго Прес (англ.), 2005. - Вип. 165. - С. 299—310. - DOI : 10.1086 / 428299 .
- ↑ А.П. Мьоллер, Дж. Ерітцое, Л.З. Гарамшегі. Коваріація між розмірами мозку та імунітетом у птахів: наслідки для еволюції розміру мозку (Англ.) // Журнал еволюційної біології (англ.): журнал. - 2005. - Вип. 18, вип. 1. - С. 223—237. - DOI : 10.1111 / j.1420-9101.2004.00805.x .
- ↑ Roth T., Sprau P., Schmidt R., Naguib M., Amrhein V. Статеві терміни пошуку матерів та розвідки території у солов’ї: нічне життя жінок (англ.) // Proc Biol Sci. : журнал. - 2009. - PMID 19324798 .
- ↑ Марлер, П. Характеристика деяких тваринних дзвінків (англ.) // Природа. - 1955. - Вип. 176. - С. 6—8. - DOI : 10.1038 / 176006a0 .
- ↑ Ленгань, Т., Дж. Лауга та Т. Оббін. Внутрішньоскладові акустичні підписи, що використовуються Королем Пінгвіном для розпізнавання батьків-пташенят: експериментальний підхід (Англ.) // Журнал експериментальної біології : журнал. - Компанія біологів (англ.), 2001. - Вип. 204. - С. 663—672.
- ↑ Торп, антихоронний спів у птахах як доказ часу реакції на пташиний слух (англ.) // Природа: журнал. - Антифональний спів у птахів як доказ часу пташиної слухової реакції. - Вип. 197. - С. 774—776. - DOI : 10.1038 / 197774a0 .
- ↑ Стокс, А., У. та Х. В. Вільямс. Антифонний дзвінок у перепелиці (англ.) // Американське орнітологічне товариство . - 1968. - Вип. 85. - С. 83—89.
- ↑ Гарріс, Тоні. Шрайкс і Буш-Шрайкс. - Університетська преса Прінстона. - С. 257–260. - ISBN ISBN 0-691-07036-9 .
- ↑ Осмастон, Б. Б. "Дует" у птахів (англ.) // Ібіс (Англ.). - Вілі-Блеквелл (англ.), 1941. - Вип. 5. - С. 310—311. - DOI : 10.1111 / j.1474-919X.1941.tb00620.x .
- ↑ Power, DM Antiphonal dueting та свідчення часу слухової реакції в помаранчевому папузі (англ.) // Американське орнітологічне товариство : журнал. - 1966. - Вип. 83. - С. 314—319.
- ↑ Хайман, Джеремі. Протидія як сигнал агресії в територіальній пісенній пташці (Англ.) // Поведінка тварин (англ.): журнал. - Ельзев'є , 2003. - Вип. 65. - С. 1179—1185. - DOI : 10.1006 / anbe.2003.2175 .
- ↑ Goodale, E. та Kotagama, SW Випробування ролі видів у зграях птахів змішаного виду дощового лісу Шрі-Ланки (англ.) // Журнал тропічної екології: журнал. - 2005. - Вип. 21. - С. 669—676. - DOI : 10.1017 / S0266467405002609 .
- ↑ Suthers Р.А. та Гектор DH Фізіологія вокалізації ехолокуючим Oilbird, Steatornis caripensis (англ.) // J. Comp. Фізіол. (англ.): журнал. - 1985. - Вип. 156, ні. 2. - С. 243—266. - DOI : 10.1007 / BF00610867 .
- ↑ Suthers Р.А. та Гектор DH Механізм для отримання ехолокаційних клацань «Сірим Свіфлом», Collocalia spodiopygia (англ.) // J. Comp. Фізіол. А (англ.): журнал. - 1982. - Вип. 148. - С. 457—470. - DOI : 10.1007 / BF00619784 .
- ↑ Коулз Р.Б., Коніші М та Петтігрю Дж. Д. Слух та ехолокація в австралійському Сірому Свіфлеті, Collocalia spodiopygia (англ.) // Журнал експериментальної біології : журнал. - Компанія біологів (англ.), 1987. - Вип. 129. - С. 365—371.
- ↑ Колліас, Н.Є. Вокальний репертуар Червоної джунглі: спектрографічна класифікація та код спілкування (англ.) // Американське орнітологічне товариство : журнал. - 1987. - Вип. 89. - С. 510—524. - DOI : 10.2307 / 1368641 .
- ↑ Еванс, CS, Македонія, Дж. М., Марлер, П. Вплив очевидних розмірів і швидкості на реакцію курей, Gallus gallus , на комп'ютерні моделювання повітряних хижаків (англ.) // Поведінка тварин (англ.): журнал. - Ельзев'є , 1993. - Вип. 46. - С. 1—11. - DOI : 10.1006 / anbe.1993.1156 .
- ↑ Пепперберг, І. М. Алекс Дослідження: когнітивні та комунікативні здібності сірих папуг. - Гарвардський університетський прес, 2000.
- ↑ Marcus, Gary F. Мова: вражаючі шпаки (англ.) // Природа. - 2006. - 27 квітня (т. 440, № 7088). - С. 1117—1118. - ISSN 0028-0836 . - DOI : 10.1038 / 4401117а .
- ↑ Янг, Джон і Гардокі, Ден. Презентація практичної майстерні з питань птахів (неопр.). Інститут регенеративного дизайну (2008). Читальний зал 7 апреля 2012 року.
- ↑ Дулінг, RJ Слухове сприйняття у птахів. Акустична комунікація у птахів, Vol. 1 (ред. DE Kroodsma & EH Miller). - 1982. - С. 95–130.
- ↑ Møller AP, J. Erritzøe, LZ Garamszegi. Коваріація між розмірами мозку та імунітетом у птахів: наслідки для еволюції розміру мозку (Англ.) // Журнал еволюційної біології (англ.): журнал. - 2005. - Вип. 18, вип. 1. - С. 223—237. - DOI : 10.1111 / j.1420-9101.2004.00805.x .
- ↑ Бонкоралліо, Г. та Нікола Сайно. Структура середовища існування та еволюція пташиної пісні: мета-аналіз доказів гіпотези про акустичну адаптацію (англ.) // Функціональна екологія: журнал. - 2007. - Вип. 21. - С. 134—142. - DOI : 10.1111 / j.1365-2435.2006.01207.x .
- ↑ Мортон Е.С. Екологічні джерела відбору на пташині звуки (англ.) // Американський натураліст (англ.): журнал. - Університет Чикаго Прес (англ.), 1975. - Вип. 109. - С. 17—34. - DOI : 10.1086 / 282971 .
- ↑ Краузе, Бернар Л. Гіпотеза про нішу (неопр.) // Звуковий вісник. - 1993. - Т. 06. Читальний зал 7 березня 2008 року. архівна копія від 7 березня 2008 на Wayback машина
- ↑ Генрік Брум. Вплив екологічного шуму на амплітуду пісні в територіальній птиці (англ.) // Журнал екології тварин : журнал. - 2004. - Вип. 73, ні. 3. - С. 434—440. - DOI : 10.1111 / j.0021-8790.2004.00814.x .
- ↑ Slabbekoorn, H. and Peet, M. Птахи співають на більш високій висоті в міському шумі (англ.) // Природа. - 2003. - Вип. 424. - С. 267. - DOI : 10.1038 / 424267а .
- ↑ 1 2 Slabbekoorn H., Smith TB Пісня, екологія та специфікація птахів (неопр.) // Philos Trans R Soc Lond B Biol Sci .. - 2002. - Т. 357, № 1420. - С. 493—503. - PMID 12028787 .
- ↑ Бейкер, МС Розпізнавання голосового діалекту та генетичні наслідки для населення (англ.) // Інтегративна та порівняльна біологія (англ.): журнал. - Oxford University Press . - Вип. 22. - С. 561—569.
- ↑ Baker, MC {{{заглавие}}} (англ.) // Бехав. Екол. Соціобіол. : журнал. - 1983. - Вип. 12. - С. 309—315.
- ↑ Chilton, G. & Lein, MR Пісня та сексуальні відповіді жіночих білокрилих горобців Zonotrichia leucophrys з популяції змішаного діалекту (англ.) // Поведінка: журнал. - 1996. - Вип. 133. - С. 173—198.
- ↑ Chilton, G., Lein, MR & Baptista, LF Mate вибір жіночих білих коронових горобців у популяції змішаного діалекту (англ.) // Бехав. Екол. Соціобіол. : журнал. - 1990. - Вип. 27. - С. 223—227.
- ↑ Мілліган, М.М. та Вернер, Дж. Дискримінація пісенного діалекту між населенням у білокрилому горобчику (англ.) // Condor: journal. - 1971. - Вип. 73. - С. 208—213.
- ↑ Beecher, MD, Campbell, SE & Nordby, JC Територіальне перебування в пісенних горобцях пов’язане з обміном піснями з сусідами, але не з розміром репертуару (англ.) // Поведінка тварин (англ.): журнал. - Ельзев'є , 2000. - Вип. 59. - С. 29—37.
- ↑ 1 2 Ratcliffe, LM & Grant, PR Визнання видів у зяблів Дарвіна (Геоспіза, Гулд). ІІІ. Відповіді чоловіків на відтворення різних типів пісень, діалектів та різновидових пісень (англ.) // Поведінка тварин (англ.): журнал. - Ельзев'є , 1985. - Вип. 33. - С. 290—307. - DOI : 10.1016 / S0003-3472 (85) 80143-3) .
- ↑ Цинк, Р. М. Генетична структура популяції та пісенні говірки у птахів (англ.) // Бехав. Мозковий Наук. (англ.): журнал. - 1985. - Вип. 8. - С. 118—119.
- ↑ Петринович, Л. Неврівноважене опитування пісенних досліджень: дим потрапляє в очі (англ.) // Бехав. Мозковий Наук. (англ.): журнал. - 1985. - Вип. 8. - С. 113—114.
- ↑ Джефрі Подос. Корельована еволюція морфології та структури голосового сигналу в зябликах Дарвіна (англ.) // Природа: журнал. - 2001. - Вип. 409. - С. 185—188. - DOI : 10.1038 / 35051570 .
- ↑ Баррінгтон, Д. Досліди та спостереження над співом птахів (англ.) // Філософські операції Лондонського королівського товариства : журнал. - 1773. - Вип. 63. - С. 249—291. - DOI : 10.1098 / rstl.1773.0031 .
- ↑ Марлер, П. та М. Тамура. Пісенні діалекти в трьох популяціях білокрилого горобця (англ.) // Кондор: журнал. - 1962. - Вип. 64. - С. 368—377. - DOI : 10.2307 / 1365545 .
- ↑ 1 2 Ноттебом, Ф. Нейронна основа пташиного польоту (англ.) // PLoS Biol : журнал. - 2005. - Вип. 3, ні. 5. - С. 163. - DOI : 10.1371 / journal.pbio.0030164 .
- ↑ Teramitsu I., Kudo LC, London SE, Geschwind DH, White SA Паралельна експресія FoxP1 і FoxP2 у мовбичці та мозку людини передбачає функціональну взаємодію (Англ.) // Й. Невроскі. (англ.): журнал. - 2004. - Вип. 24, ні. 13. - С. 3152—3163. - DOI : 10.1523 / JNEUROSCI.5589-03.2004 . - PMID 15056695 .
- ↑ Nottebohm, F. Дорога, якою ми пройшли: відкриття, хореографія та значення нейронів, що змінюються на мозку (англ.) // Енн. NY Акад. Наук. (англ.): журнал. - 2004. - Вип. 1016 рік. - С. 628—658. - DOI : 10.1196 / літописи.1298.027 .
- ↑ Бреновіц, Еліот А. та Майкл Д. Бічер. Пісенне навчання у птахів: різноманітність та пластичність, можливості та завдання (Англ.) // Тенденції нейронаук (англ.): журнал. - Клітинний прес (англ.), 2005. - Вип. 28, ні. 3. - С. 127—132. - DOI : 10.1016 / j.tins.2005.01.004 .
- ↑ Слейтер, PJB Пісенне навчання пісні: причини та наслідки (англ.) // Етол. Екол. Evol. (англ.): журнал. - 1989. - Вип. 1. - С. 19—46.
- ↑ Thorpe, W. Процес пісенного навчання в полоні, що вивчається за допомогою звукового спектографа (англ.) // Природа: журнал. - 1954. - Вип. 173. - С. 465—469. - DOI : 10.1038 / 173465a0 .
- ↑ Коніші, М. Роль слухового зворотного зв’язку щодо контролю за вокалізацією у білоголового горобця (англ.) // Zeitschrift fur Tierpsychologie: journal. - 1965. - Вип. 22. - С. 770—783.
- ↑ Мова птахів: Вокальне привласнення (неопр.). Слухайте природу . Британська бібліотека. Читальний зал 7 апреля 2012 року.
- ↑ Мова птахів: «Розмовні» птахи (неопр.). Слухайте природу . Британська бібліотека. Читальний зал 7 апреля 2012 року.
- ↑ Мова птахів: називає хибною мімікою (неопр.). Слухайте природу . Британська бібліотека. Читальний зал 7 апреля 2012 року.
- ↑ Брайнард, М. С. та Дупе, AJ Слухові відгуки у навчанні та підтримці голосової поведінки (англ.) // Nature Rev. Neurosci. : журнал. - 2000. - Вип. 1. - С. 31—40. - DOI : 10.1038 / 35036205 .
- ↑ Кері, Томас Дж. Поведінкова нейробіологія: клітинна організація природної поведінки. - Sinauer Associates, Inc., 2000. - ISBN ISBN 978-0-87893-092-0 .